El camino del agua
Código 03A8 Letra griega PSI Ψ
El Camino del agua
Movimientos del agua a través del xilema
Dada la importancia del agua en las plantas, hemos querido profundizar un poco en el conocimiento de los procesos que se relacionan con ella. Independientemente de sus usos en bonsái, este artículo quiere exponer las bases técnicas de cómo se producen estos procesos.
Propiedades físico- químicas del agua
El agua es una molécula simple de bajo peso molecular con un anómalo comportamiento físico-químico derivado de su estructura molecular, lo que la convierte en una sustancia diferente a la mayoría de los líquidos.
La molécula de agua está formada por dos átomos de H unidos a un átomo de O por medio de dos enlaces covalentes parcialmente iónicos (2/3 carácter covalente y 1/3 carácter iónico).
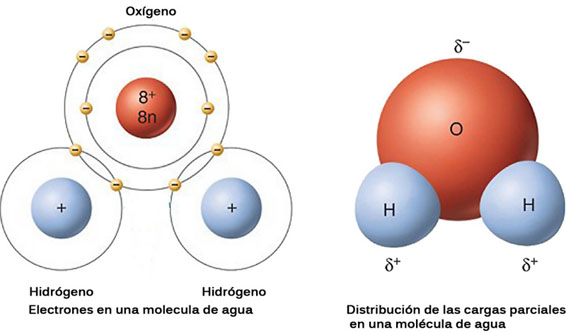
A temperatura ambiente es líquida, al contrario de lo que cabría esperar, ya que otras moléculas de parecido peso molecular (SO2, CO2, SO2, H2S, etc) son gases. Este comportamiento se debe a que los dos electrones de los dos hidrógenos están desplazados hacia el átomo de oxigeno, por lo que en la molécula aparece un polo negativo, donde está el oxígeno, debido a la mayor densidad electrónica, y dos polos positivos, donde están los dos hidrógenos, debido a la menor densidad electrónica. La molécula de agua es un dipolo.
Entre los dipolos del agua se establecen fuerzas de atracción llamados puentes de hidrógeno, formándose grupos de 3-9 moléculas. Con ello se consiguen pesos moleculares elevados y el agua se comporta como un líquido. Estas agrupaciones, le confieren al agua sus propiedades de fluido, en realidad, coexisten estos pequeños polímeros de agua con moléculas aisladas que rellenan los huecos.
Los enlaces por puentes de hidrógeno son, aproximadamente, 1/20 más débiles que los enlaces covalentes, el hecho de que alrededor de cada molécula de agua se dispongan otras moléculas unidas por puentes de hidrógeno, permite que se forme en el seno del agua una estructura ordenada de tipo reticular, responsable en gran parte del comportamiento anómalo y de sus propiedades físicas y químicas.
Las fuerzas de cohesión que se establecen entre las moléculas superficiales son diferentes a las del interior. Mientras las moléculas bajo la superficie líquidas experimentan fuerzas de atracción con otras moléculas vecinas en todas las direcciones, las que se encuentran en la superficie están ligadas sólo por otras moléculas superficiales y por aquellas ubicadas inmediatamente debajo. Esto crea una mayor tensión sobre la superficie del líquido, llamada “tensión superficial”.
La tensión superficial, no solo influye en la superficie del agua o en la forma de las gotas, sino que puede aportar presión al resto del líquido. Como veremos más adelante, la tensión superficial en las superficies de evaporación de las hojas, generará la fuerza física para elevar el agua a través del sistema vascular de la planta.
Los enlaces de hidrógeno en el agua también aportan otra característica fundamental denominada “cohesión”, basada en la mutua atracción de las moléculas de agua. Otra propiedad relacionada con la anterior es la “adhesión”, y es la atracción del agua a un medio sólido, como puede ser la pared celular o una superficie de cristal. Cohesión, adhesión, y tensión superficial unidos nos aportan otro fenómeno conocido como “capilaridad”. Es el movimiento del agua a través un tubo capilar.
Si disponemos un fino tubo de cristal en vertical, el movimiento ascendente del agua es debido a la atracción del agua a la superficie del tubo de cristal (adhesión) y a la tensión superficial del agua, que tiende a minimizar el área expuesta en la combinación agua-aire. Juntos, adhesión y tensión superficial elevan las moléculas de agua en el tubo hasta que la fuerza ascendente es equilibrada por el peso de la columna de agua. Cuando más fino sea el tubo, mayor altura alcanza el agua.
Transpiración
Podríamos definir la transpiración como el movimiento del agua a través de la planta, desde la raíz hasta las hojas durante el proceso de fotosíntesis. Todos los procesos son necesarios, y no ocurren uno sin el otro. Por ejemplo, no habrá circulación ascendente de agua si no hay fotosíntesis, del mismo modo que no habrá fotosíntesis sin un sistema radicular capaz de proporcionar el agua necesaria.
![]()
Ejemplos de transpiración
Una sola planta de maíz, es capaz de evaporar 196 litros de agua en los cuatro meses que dura su cultivo.
Para la producción de un kg de materia seca, son necesarios entre 200 y 900 litros de agua
Vías de la transpiración
Agua en el suelo disponible para las plantas es únicamente la que se encuentra en forma líquida, las raíces no pueden absorber el agua que forma el vapor contenido en el aire entre las partículas de suelo
Las raíces absorben el agua y ésta pasa a través de la epidermis de la raíz al córtex. Desde el córtex, continúa hacia la endodermis. La endodermis controla la absorción de solutos y envía el agua y las sales disueltas en ella hacia el xilema
El agua es elevada a través del xilema hasta el xilema de la hoja, y desde allí se distribuye entre el mesófilo
Por último, el agua acaba evaporada en la cámara estomática, desde donde es dispersada a la atmósfera siempre y cuando los estomas se encuentren abiertos.
Transporte del agua a través del xilema
Potencial hídrico
El agua en estado líquido es un fluido cuyas moléculas se hallan en constante movimiento. La movilidad de estas moléculas dependerá de su energía libre, es decir de la fracción de la energía total que puede transformarse en trabajo. La magnitud más empleada para expresar y medir su estado de energía libre es el potencial hídrico (Ψw). El Ψw se mide en atmósferas, bares, y megapascales (Mpa), siendo 0,987 atm = 1 bar = 0,1 Mpa, y suele ser representado por la letra griega “PSI” Ψ. A una masa de agua pura, libre, sin interacciones con otros cuerpos, y a presión normal, le corresponde un Ψw igual a 0.
El Ψw está fundamentalmente determinado por la presión y por la actividad del agua. Esta última depende, a su vez, del efecto osmótico, presencia de solutos, y del efecto de la gravedad.
| Potencial hídrico y sus componentes (en Mpa) | ||||
| Potencial hídrico | Presión hidrostática | Potencial osmótico | Gravedad | |
| a- Aire exterior (humedad relativa = 50%) | -95.2 | |||
| b- Aire interior de la hoja | -0.8 | |||
| c- Pared celular del mesófilo | -0.8 | -0.7 | -0.2 | 0.1 |
| d- Vacuola del mesófilo | -0.8 | 0.2 | -1.1 | 0.1 |
| e- Xilema de la hoja | -0.8 | -0.8 | -0.1 | 0.1 |
| f- Xilema de la raíz | -0.6 | -0.5 | -0.1 | 0.0 |
| g- Vacuola de la célula de raíz | -0.6 | 0.5 | -1.1 | 0.0 |
| h- Suelo adyacente a la raíz | -0.5 | -0.4 | -0.1 | 0.0 |
| i- Suelo a 10mm de la raíz | -0.5 | -0.2 | -0.1 | 0.0 |
El Ψw se puede expresar en la suma de sus componentes:
Ψ w = Ψp + Ψs + Ψg
El Ψp, presión hidrostática, es nulo a presión atmosférica, positivo para presiones por encima de la atmosférica, y negativo en condiciones de tensión o vacío.
El Ψs, potencial osmótico, representa la disminución de la capacidad de desplazamiento del agua debido a la presencia de solutos. A medida que la concentración de soluto (es decir, el número de partículas de soluto por unidad de volumen de la disolución) aumenta, el Ψs se hace más negativo. Sin la presencia de otros factores que alteren el potencial hídrico, las moléculas de agua de las disoluciones se moverán desde lugares con poca concentración de solutos a lugares con mayor concentración de soluto.
El Ψs se considera 0 para el agua pura.
El Ψg, potencial gravitacional, representa la fuerza ejercida sobre el agua por la gravedad. A mayor altura del suelo, mayor Ψg
Teoría de la tensión-cohesión
En teoría, la presión necesaria para mover el agua a través del xilema se crea sumando la presión positiva generada por las raíces, y la presión negativa creada en las hojas.
Cabe considerar la presión generada por la raíz como insignificante, ya que generalmente suele ser inferior a 0.1MPa. En cambio, las hojas al transpirar desarrollan una gran tensión (presión hidrostática negativa –Ψp), y ésta es la presión que “estira” el agua hacia arriba a través del xilema. Este mecanismo es conocido como “teoría de la tensión-cohesión” ya que requiere de las propiedades cohesivas del agua para mantener grandes tensiones en la columna de agua del xilema.
El potencial hídrico llega a ser enorme en árboles grandes, alcanzando en algunos casos las 30 atmósferas.
Cuando la tensión llega al máximo las propiedades cohesivas del agua no son suficientes para mantener unida la columna de agua, entonces ésta se rompe, generando una burbuja de aire en el xilema.
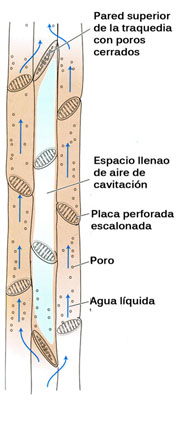
Este efecto se denomina cavitación, y es solventado por la planta derivando el flujo de agua a los canales contiguos (traqueidas). De este modo, el árbol evita que la cavitación bloquee totalmente el flujo de agua ascendente.
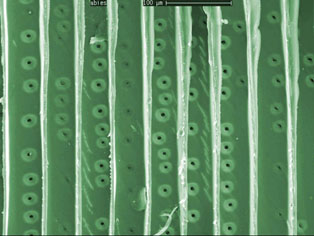
Traqueidas de Picea sp. mostrando las punteaduras porosas entre traqueidas
Las traqueidas están formadas por las paredes de células muertas elongadas en forma de huso y dispuestas en filas verticales, formando vasos cerrados con membranas oblicuas con las que se unen de forma vertical. El agua puede pasar de unas traquedias a otras a través de las membranas superiores, o a través de las paredes laterales por medio de unos pequeñas punteaduras microscópicas que comunican una traqueida con otra mediante un pequeño poro que actúa a modo de válvula. Es poroso a muy baja presión, pero posee la capacidad de bloquearse al aumentar esta presión, aislando de este modo las peligrosas burbujas de aire producidas por la cavitación.
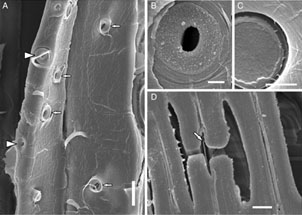
Pié de foto: A Punteaduras mostrando los poros entre traqueidas. B y C poros de maderas vieja y mueva. D corte trasversal de una punteadura, mostrando la estructura que hace las funciones de válvula.
En condiciones normales, al reducirse la tensión (ciclo nocturno), las burbujas de gas son reabsorbidas, y el flujo restaurado.
Presión radicular
Como ya hemos mencionado anteriormente, la raíz genera una pequeña presión positiva que empuja el agua hacia arriba. Tan pequeña, que no afecta en lo más mínimo al movimiento del agua en la planta, pero en cambio sí tiene la importancia de iniciar el movimiento desde el suelo hacia las vías interiores de distribución.
El potencial hídrico del suelo puede ser dividido en dos componentes: el potencial osmótico Ψs, y la presión hidrostática Ψp
El agua se mueve en el suelo, si exceptuamos la gravedad, predominantemente por diferencias entre el potencial osmótico. Al ser el potencial osmótico en el interior de la raíz menor que el del agua en contacto con ella, se genera un flujo a causa de esta diferencia de potencial hídrico. Y al absorber las plantas el agua más cercana a la raíz, se produce una disminución de la presión hidrostática Ψp alrededor de los pelos capilares y de la epidermis de la raíz. Esto provoca una diferencia de presión en las zonas vecinas, creando un flujo hacia las áreas donde el agua ha sido absorbida.
En suelos muy secos, la presión hidrostática Ψp puede caer por debajo del denominado “punto de marchitez”. En este punto, el potencial hídrico del suelo es tan bajo que las plantas no son capaces de generar ningún flujo de agua alrededor de la raíz, mostrando síntomas de marchitez debido a la imposibilidad de reponer el agua perdida por la traspiración. Esto significa que el potencial hídrico total del suelo Ψw, es menor o igual al potencial osmótico de la planta Ψs.
Esto mismo sucede cuando la concentración de sales en el suelo es muy alta debido al abonado, creando un suelo hipertónico. El potencial osmótico del suelo será menor al interior de la raíz, impidiendo así, la absorción del agua por ésta. Los síntomas producidos por exceso de sales, se asemejan a los de la sequía
Absorción
El contacto íntimo entre la superficie de la raíz y el suelo es esencial para una efectiva absorción de agua. Los “pelos radicales” son microscópicas extensiones de células epidérmicas que aumentan considerablemente la superficie de absorción. En plantas adultas y en época de crecimiento, estos pelos suponen más del 60% de la superficie de la raíz.
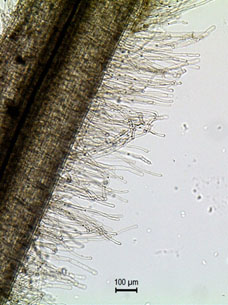
La absorción se produce únicamente en las zonas apicales de la raíz, y a través de los pelos radicales. Las raíces maduras son incapaces de absorber agua del suelo.
Viendo la importancia de los pelos radicales en la absorción del agua, es fácil entender los problemas de enraizado. Al manipular el suelo, la mayoría de estos microscópicos pelos, frágiles como cristal, son desprendidos, imposibilitando así que la planta pueda reponer el agua transpirada.
La raíz en crecimiento, solo se desarrolla en suelo húmedo. Si entra en contacto con un área con presión hidrostática muy baja, detendrá su crecimiento.
